Conformational dynamics of the inner pore helix of voltage-gated potassium channels
Abstract
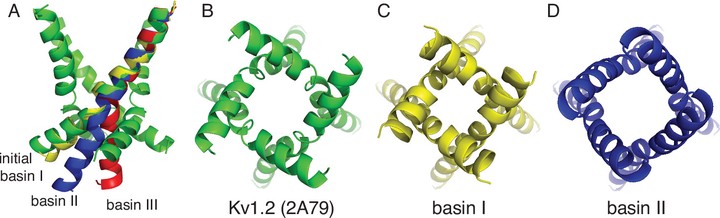
Voltage-gated potassium (Kv) channels control the electrical excitability of neurons and muscles. Despite this key role, how these channels open and close or gate is not fully understood. Gating is usually attributed to the bending and straightening of pore-lining helices at glycine and proline residues. In this work we focused on the role of proline in the Pro-Val-Pro (PVP) motif of the inner S6 helix in the Kv1.2 channel. We started by developing a simple hinged-rod model to fully explore the configurational space of bent helices and we related these configurations to the degree of pore opening. We then carried out fully atomistic simulations of the S6 helices and compared these simulations to the hinged-rod model. Both methods suggest that Kv1 channels are not tightly closed when the inner helices are straight, unlike what is seen in the non-PVP containing channels KcsA and KirBac. These results invite the possibility that the S6 helices may be kinked when Kv1 channels are closed. Our simulations indicate that the wild-type helix adopts multiple spatially distinct configurations, which is consistent with its role in adopting a closed state and an open state. The two most dominant configurational basins correspond to a 6 Å movement of the helix tail accompanied by the PVP region undergoing a local α-helix to 310-helix transition. We explored how single point mutations affect the propensity of the S6 helix to adopt particular configurations. Interestingly, mutating the first proline, P405 (P473 in Shaker), to alanine completely removed the bistable nature of the S6 helix possibly explaining why this mutation compromises the channel. Next, we considered four other mutations in the area known to affect channel gating and we saw similarly dramatic changes to the helix’s dynamics and range of motion. Our results suggest a possible mechanism of helix pore closure and they suggest differences in the closed state of glycine-only channels, like KcsA, and PVP containing channels.